Life at the Cell and Below-Cell Level. The Hidden History of a Fundamental Revolution in Biology
"Dr. Ling is one of the most inventive biochemist I have ever met." |
Chapter 11. The Polarized Multilayer Theory of Cell Water (p. 74-108) |
|
11.1 Background Before presenting the polarized multilayer (PM) theory, it behooves one to examine earlier ideas of water structuring around proteins and other macromolecules (for a more extensive review, see Reference 155). Briefly in 1953, in more detail in 1955, В. Jacobson offered a "water structure
hypothesis" to explain the dielectric behaviors of aqueous macromolecular
solutions. He said: "If the macromolecule has many oxygen and nitrogen
atoms on the surface in such positions that they fit into the ideal water
lattice, a very pronounced ordering effect is obtained and result in an almost
ideal four-coordinated structure."334 p 2923;
384 Jacobson described the hydration shells thus formed as
"ice-like" to distinguish it from ordinary liquid water, which also
has a four-coordinated structure385; 386 but not nearly as
perfect. Two years later, Nobel Laureate, Albert Szent-Györgyi took the notion of ice-like water one
step further. In Szent-Györgyi's "iceberg
theory," crystalline ice surrounds protein molecules in living
cells.387 pp 32-37 Ice-like hydration water on proteins was also
advocated by Klotz.388 However, there is serious contradictory evidence against the
iceberg theories. 11.2 Polarized multilayer theory of cell water and its world-wide
confirmation The polarized multilayer (PM) theory of
cell water, an integral part of the association-induction (AI) hypothesis,154;
15 pp 271-310; 107 pp 69-110 was published three years after the
publication of the AI Hypothesis (AIH) proper. Notwithstanding, I have chosen
to present the PM theory here first and the AIH proper later, because like
LFCH, the PM theory also deals with the static aspects of the living
cell. In contrast, the dynamic or inductive aspect—built upon the
foundation of the static aspects—will be presented as part of the AI
Hypothesis proper later. In the PM theory all or virtually all the water in
resting living cells assumes a dynamic structure different from normal liquid
water, owing primarily to its interaction with a matrix of
"fully-extended" protein chains present in all living cells. By fully-extended, I mean that all the backbone
NH and CO groups of the protein are not engaged in inter- or
intra-macromolecular H-bonds. The fully-extended conformation is not to be
confused with the "extended" conformation, which is sometimes used to
denote the "β-pleated sheet" conformation.485 p 501 Under
certain conditions, the fully-extended conformation corresponds to what is
called "random coil" and what Bungenberg
de Jong called linear conformation [6.2(1)].
But according to the PM theory, the fully-extended proteins in living cells
and the water they dominate are not randomly distributed [15.1(1)]. Fully-extended protein chains carry on their
polypeptide chains or backbones a repeating sequence of properly-spaced
negatively charged CO (referred to as N sites) and positively charged NH groups
(referred to as P sites), both directly exposed to the bulk-phase cell water.
Such a matrix of fully-extended proteins chains constitutes an NP-NP-NP system.155
A two-dimensional checkerboard of N and P sites
represents an NP system. NO-NO-NO system refers to chains carrying
properly-spaced N sites alternating with vacant 0 sites. Conversely, a
PO-PO-PO system refers to chains carrying properly-spaced P sites alternating
with vacant 0 sites.
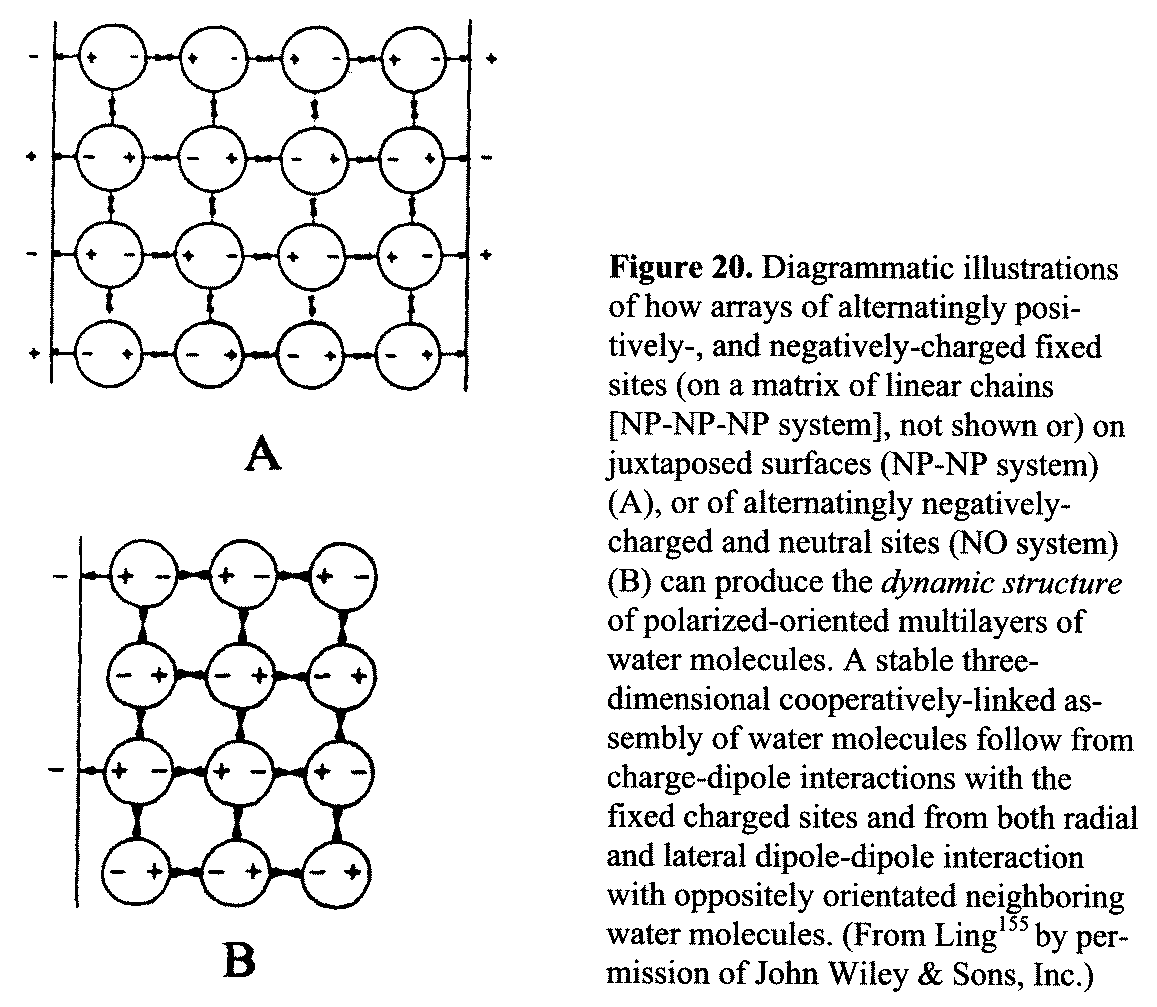
Figure 20. Diagrammatic illustrations of
how arrays of alternatingly positively-, and negatively-charged fixed sites (on
a matrix of linear chains [NP-NP-NP system], not shown or) on juxtaposed
surfaces (NP-NP system) (A), or of alternatingly negatively-charged and neutral
sites (NO system) (B) can produce the dynamic structure of polarized-oriented
multilayers of water molecules. A stable three-dimensional
cooperatively-linked assembly of water molecules follow from
charge-dipole interactions with the fixed charged sites and from both radial
and lateral dipole-dipole interaction with oppositely orientated neighboring
water molecules. (From Ling155 by permission of
John Wiley & Sons, Inc.) Note that having a vacant site and having
no site between each pair of nearest neighboring N or P sites are not the
same, owing to the different distances separating the nearest-neighboring N or P sites.
Now, water molecules are special because
the two positively-charged hydrogen atoms are not symmetrically placed on
either side of the negatively-charged oxygen atom, but are asymmetrically
placed, not unlike those shown in the formula, H2O. Accordingly,
each water molecule may be seen as a dipole with a positive end and a
negative end and a well-defined (permanent) dipole moment, equal to 1.86 debye. On the properly-spaced, alternating N and P sites of a
fully-extended protein chain are oriented and polarized a layer of
oppositely-oriented water dipoles. This first layer of polarized-oriented,
adsorbed water molecules in turn adsorbs, orients and
polarizes a second layer of oppositely-oriented water dipoles. And this
continues until a dynamic structure of multiple layers
of polarized and oriented water are built up. Figure 20, intended to
illustrate this dynamic structure, is unfortunately itself a static picture;
and, as such, exaggerates the orderliness and stability of the dynamic water
structure.
To be continued |
|
|
Разделы книги
Contents (PDF
218 Kb) |
|
|
На страницу
книги "Life at the Cell and Below-Cell Level..." |
|

{kind=link}